编辑|易霄寒
非生物胁迫是环境因素中对农作物影响最为广泛的胁迫因子,统计数据表明,1964—2007年间,全球作物(水稻、玉米和小麦)产量,因高温和干旱导致的损失高达9%~10%,在某些区域损失更高。
其中,热胁迫能影响作物的多个生理生化过程,如高温条件下,玉米胚芽鞘的生长发育减缓,叶片光合效率降低,玉米雄穗分枝以及雌花小花密度明显减少,籽粒产量降低,以及玉米籽粒中淀粉、蛋白质和油脂等成分的含量下降等。
实验室前期研究表明,ZmNF-YC13基因受内质网胁迫诱导表达,在玉米中超表达ZmNF-YC13可提高植株对内质网胁迫的耐受性。
本研究表明ZmNF-YC13还受热胁迫和渗透胁迫快速诱导表达,通过热诱导表达启动子驱动ZmNF-YC13基因,可显著提高植株对高温的耐受性。
表达分析等表明ZmNF-YC13在高温条件下,可能是通过增强热胁迫应答基因的表达来提高植株的耐热能力。
本研究不仅丰富了NF-YC转录因子在热胁迫应答通路中的调控模式,也为利用该位点进行耐热性辅助选择和种质资源鉴定,提供了理论依据。
实验室苗期玉米材料均播于Pindstrup基质中,置于光照培养箱培养,参数设置为温度28/25℃,光周期为16h/8h,光照强度250µmolm-2s-1,湿度40%~60%。
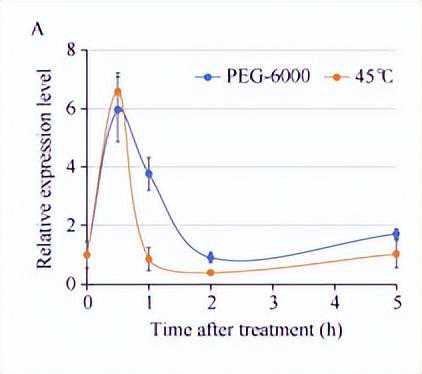
对ZmNF-YC13基因在高温处理下的表达模式进行分析时,转基因受体材料播种后第10天,保留长势一致的植株并提前一天浇足水,将培养箱温度调为45℃光照强度125µmolm-2s-1,分别在0、0.5、1、2和5h取第2叶相同部位组织,液氮速冻后于超低温冰箱保存待用。
对ZmNF-YC13基因在渗透胁迫处理下的表达模式进行分析时,播种后第10d,保留长势一致的植株,用10%的PEG-6000溶液彻底浇透,分别在0、0.5、1、2和5h取第2叶相同部位组织,液氮速冻后于超低温冰箱保存待用。
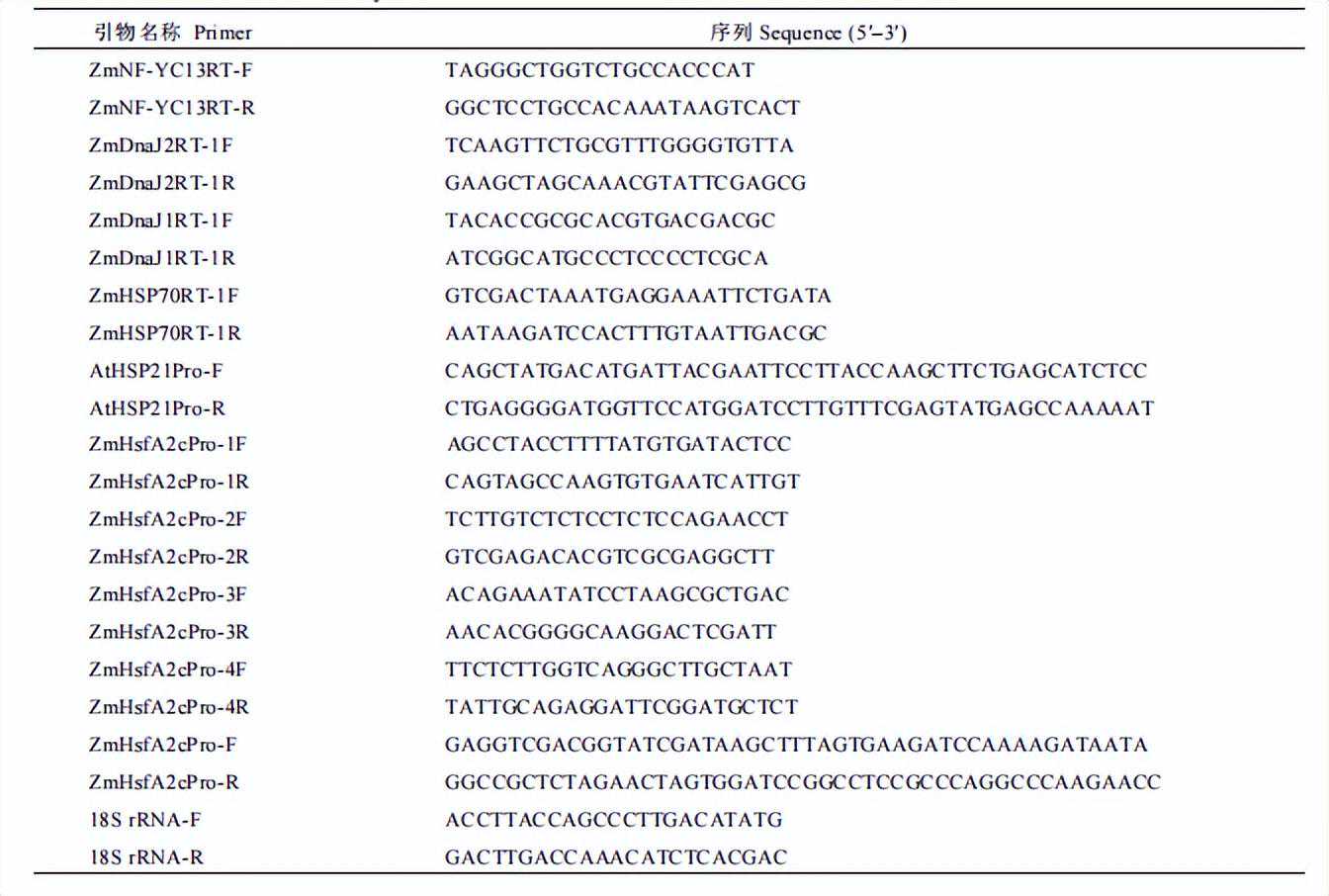
利用热敏感和耐热自交系进行表达分析时,在播种后10d,保留长势一致的植株,并提前一天浇足水,将培养箱温度调为45℃光照强度125µmolm-2s-1,分别在0h和0-5h取第2叶相同部位组织,液氮速冻后于超低温冰箱保存待用。
每个样品设置3个技术重复,18SrRNA用作内参基因,利用比较CT值法进行计算相对表达量。
首先实验将分析玉米苗期高温处理及表型,我们选择籽粒大小接近的野生型和ZmNF-YC13热诱导表达材料玉米种子播于Pindstrup基质中,置于光照培养箱培养,温度28/℃25,℃光周期16h/8h,光照强度250µmolm-2s-1,湿度40%~60%。
播种后第10天,保留长势一致的植株并提前一天浇足水,将培养箱温度调为45,℃光照强度125µmolm-2s-1,继续培养8h。
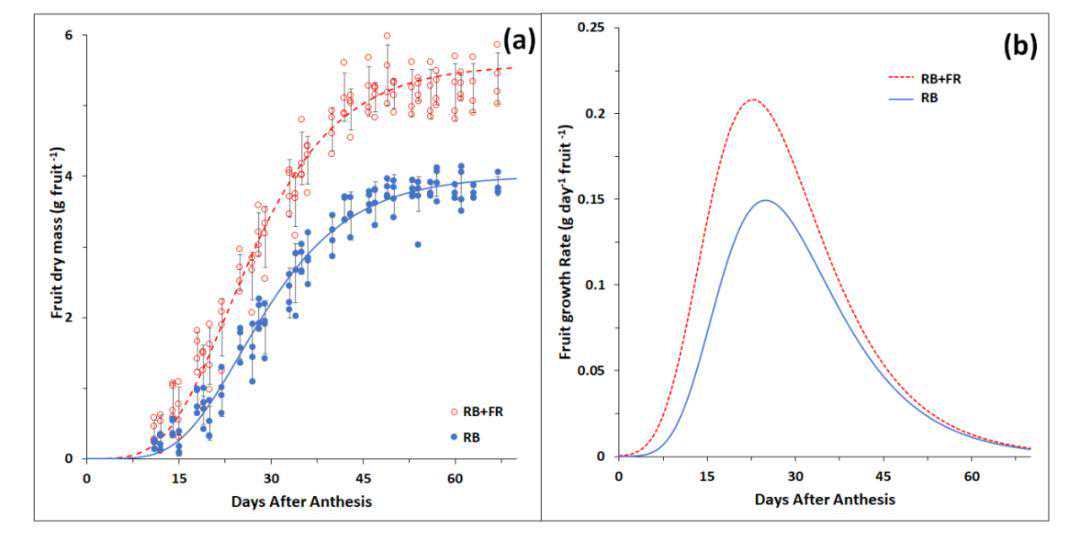
处理结束后将培养箱参数调至正常模式继续培养15d,培养过程中适时补充水分和营养。
恢复培养结束后,分别测量第5叶和第6叶(心叶)长、第5叶宽、地上部粗(地下与地上连接处以上1cm位置,用游标卡尺测量)和地上部分、地下部分鲜重。
测干重前,先用110℃烘6h,再用80℃烘干至恒重。
接着将生长14d的HSP21Pro,ZmNF-YC13-myc植株叶片(45℃处理2h后)切成0.5~1.0cm条状,然后放入含1%甲醛的固定液中,冰上真空(0.08MPa)交联10min后,加入甘氨酸(终浓度为0.125molL–1),再真空渗透10min,停止交联,ddH2O冲洗3次,用滤纸快速吸干材料表面液体后放入液氮速冻。
琼脂糖磁珠用于细胞裂解液预清除,GFP-Trap琼脂糖磁珠和Myc-Trap琼脂糖磁珠用于免疫沉淀。qPCR检测分析后,以GFP-Trap作为对照进行计算富集度,分别进行3次独立实验。
在实验室前期研究中,筛选出了响应内质网胁迫应答调控因子ZmNF-YC13,并证实该基因在玉米内质网胁迫应答调控过程中具有重要作用。
进一步对该基因在其他非生物胁迫条件下的分析表明,ZmNF-YC13基因的表达受到渗透胁迫和高温的快速诱导,随后下降。
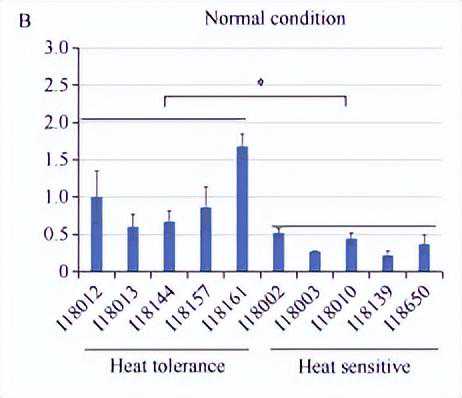
为进一步分析ZmNF-YC13基因表达量与玉米耐热性的关系,我们选取了在耐热能力上具有极端差异(耐热型和热敏感型)的玉米自交系,并于高温胁迫处理前后分别检测了ZmNF-YC13基因的表达量。
结果表明,在正常条件下,耐热型自交系材料中ZmNF-YC13基因的表达量,均高于热敏感型自交系材料。
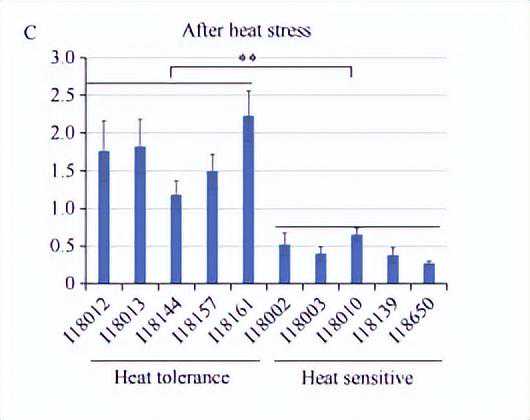
在45℃高温处理1h后,ZmNF-YC13基因在各自交系材料中出现了不同程度的响应,在耐热型自交系材料中上调水平更高,且该基因整体的表达水平高于热敏感型自交系材料。
上述结果说明,ZmNF-YC13基因可能参与玉米热胁迫应答调控过程。
组成型表达ZmNF-YC13虽在一定程度上提高玉米植株对非生物胁迫的抗性,但过高的ZmNF-YC13基因表达量同时也会抑制植株的生长。
为避免提高ZmNF-YC13基因表达量对植株生长带来的迟缓效应,我们克隆了拟南芥热诱导表达基因AtHSP21基因启动子,构建了利用该启动子表达ZmNF-YC13基因的玉米转基因材料(HSP21Pro:ZmNF-YC13-myc)。
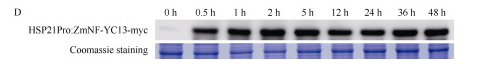
随后我们在高温处理前以及高温处理后不同时间段利用anti-myc抗体在蛋白水平上对T3代纯合材料HSP21Pro:ZmNF-YC13-myc。
检测表明,在正常培养条件下(0h),ZmNF-YC13-myc融合蛋白的表达量较低;受到45℃高温处理后,ZmNF-YC13-myc的蛋白水平快速积累。
该结果表明成功构建了ZmNF-YC13基因的热诱导表达玉米材料。

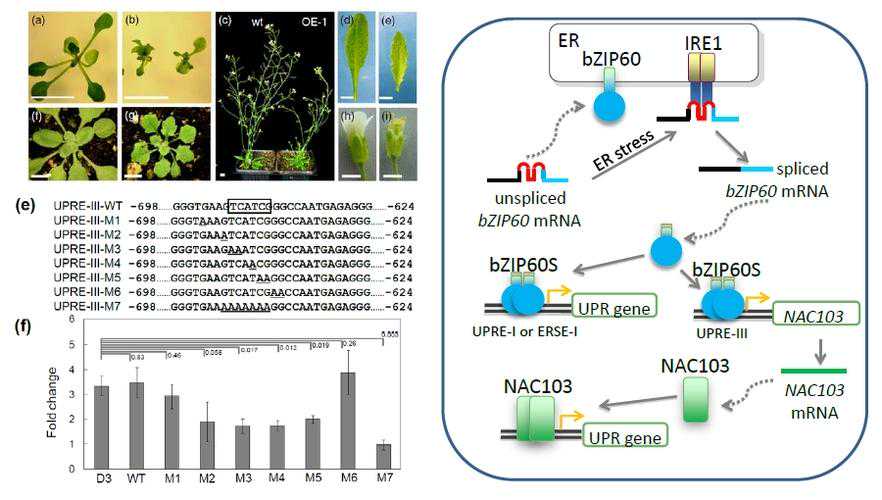
我们进一步对通过热诱导表达ZmNF-YC13基因是否能够提高玉米苗期植株的耐热能力进行了分析。
在正常培养条件下,野生型植株与ZmNF-YC13热诱导表达植株的长势无显著差异。

在经过45℃高温胁迫处理8h后,野生型植株和ZmNF-YC13热诱导表达植株的叶片,均出现了不同程度的卷曲和萎蔫现象。

将处理后的植株在正常条件下恢复培养15d后,ZmNF-YC13热诱导表达植株的长势均优于野生型的植株。
同时,我们对野生型植株和ZmNF-YC13热诱导表达植株,在正常培养条件下和在高温处理后的生物量进行了测定。

结果表明,在高温处理之后,ZmNF-YC13热诱导表达植株的地上部和地下部的鲜重和干重均显著高于野生型植株;第6叶、第5叶长度和宽度以及地上部粗度(地下部和地上部连接处以上1cm位置)均显著高于野生型植株。
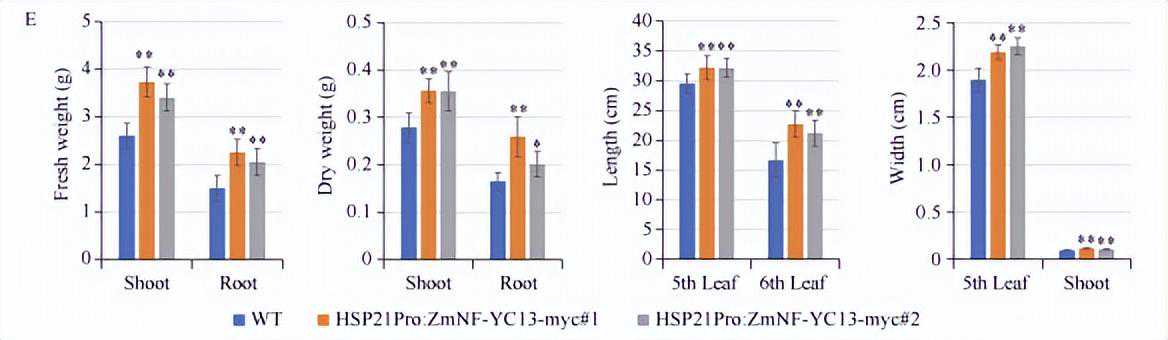
以上结果说明,热诱导表达ZmNF-YC13基因能够提高玉米苗期植株的耐热能力。
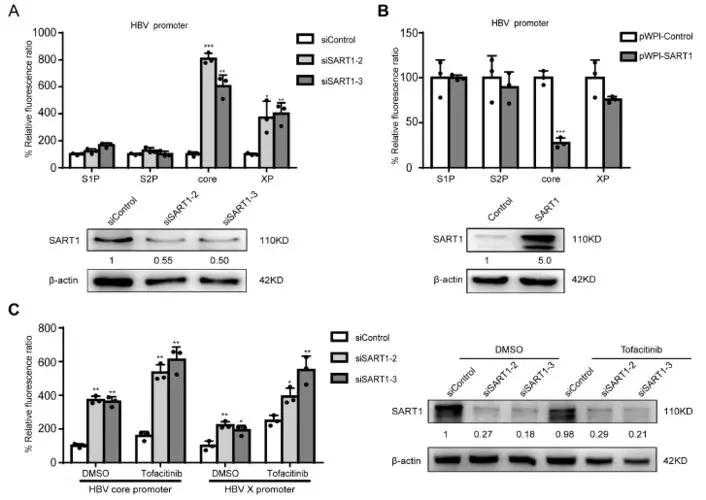
而由于高温胁迫是自然条件下引起内质网胁迫的主要因素之一,因此我们猜测ZmNF-YC13在内质网胁迫应答通路中调控的下游基因(ZmDnaJ1、ZmDnaJ2、ZmHSP70和ZmHsfA2c等)可能同样参与了热胁迫应答调控。
为证实这一猜测,我们首先对受ZmNF-YC13调控的下游基因,在高温处理下的表达特点,进行了分析。
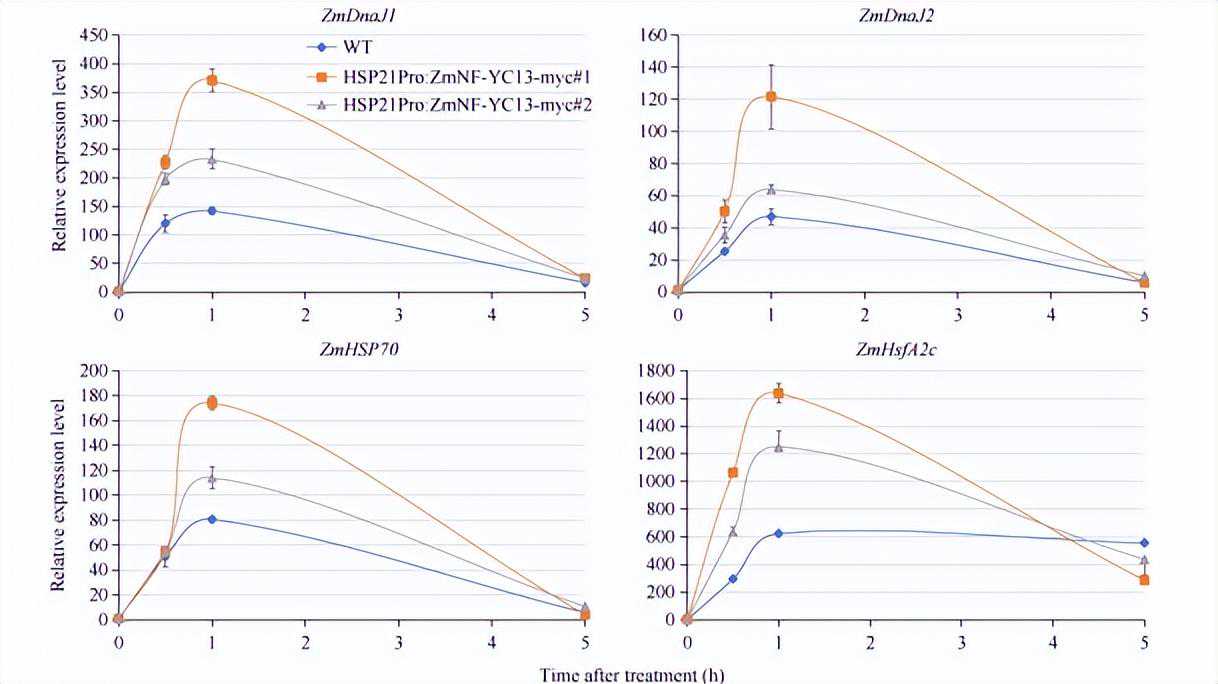
结果表明,在45℃高温处理下,野生型玉米植株中ZmDnaJ1、ZmDnaJ2、ZmHSP70和ZmHsfA2c基因的表达量快速增加,且在1h左右达到高峰,说明ZmDnaJ1、ZmDnaJ2、ZmHSP70和ZmHsfA2c基因是热胁迫应答基因。
在ZmNF-YC13热诱导表达材料中,ZmDnaJ1、ZmDnaJ2、ZmHSP70和ZmHsfA2c基因受高温诱导表达的程度较野生型植株明显提高,表明提高ZmNF-YC13基因表达量能增强热胁迫诱导表达基因的表达。
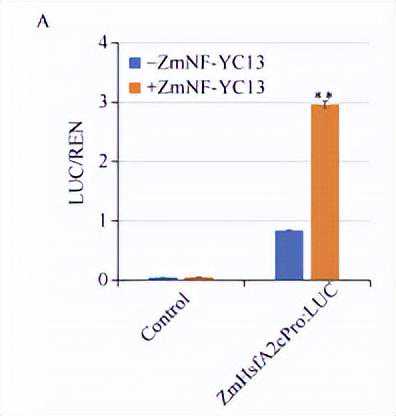
由于热应激转录因子在植物热胁迫应答调控中具有重要作用,且Gu等研究表明在拟南芥中超表达HSFA2(ZmHsfA2c)可提高植株耐热能力,而在本研究中发现ZmHsfA2c的表达量受ZmNF-YC13的调控。
为了进一步证实ZmNF-YC13-ZmHsfA2c这一调控通路,我们利用荧光素酶报告基因实验和ChIP-qPCR进一步进行了分析。
荧光素酶报告基因实验结果表明,ZmNF-YC13蛋白可显著激活ZmHsfA2c基因的启动子。
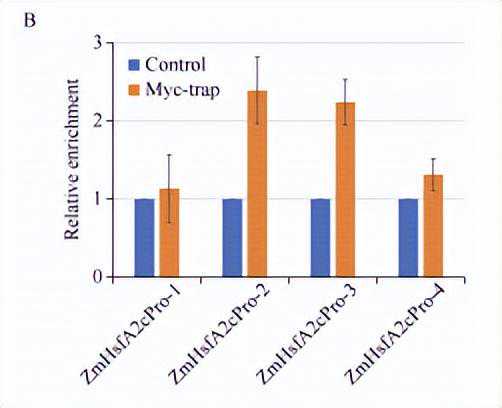
利用染色质免疫共沉淀并结合qPCR的方法,对ZmHsfA2c基因起始密码子ATG上游1kb范围的启动子区域分析表明,目标蛋白在ZmHsfA2c基因的启动子区段2和区段3上有较高的富集峰。
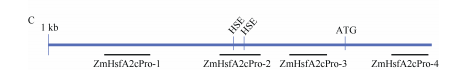
我们进一步对启动子区段2和区段3的序列分析发现,在区段中存在两个典型的热胁迫应答元件(heatshockelement,HSE)。
以上结果表明,ZmNF-YC13能够调控ZmHsfA2c基因表达,可能依赖于热胁迫应答元件。
植物作为一种固着生物,其生长发育极易受到自然环境的影响,如高温破坏植物细胞膜系统的稳定性、引起细胞内酶系统紊乱、籽粒过早脱水等,从而扰乱其正常的生理阶段和进程,使生长发育和产量形成受到严重影响。
植物在进化过程中从分子、细胞和生理生化层面上也形成了一系列的调节机制,以应对或缓解环境变化所带来的影响。
如在高温胁迫下,植物细胞首先感知高温刺激并将信号放大传递至胞内,进而激活一系列高温胁迫应答基因的表达,以此来应对或缓解高温带来的损伤,这一信号转导机制和转录调控网络在植物响应高温胁迫过程中至关重要。
而与热胁迫应答相关的转录因子,作为调控下游靶基因表达的分子开关,在这个过程中具有不可替代的作用。
Mei等研究发现ZmNF-YC13基因受内质网胁迫(用化学试剂二硫苏糖醇和衣霉素处理)诱导表达,编码核内蛋白,且具备一定的转录激活能力;在玉米中组成型表达ZmNF-YC13基因后,能够提高植株对内质网胁迫的耐受能力。
在自然环境中,高温是诱导产生内质网胁迫的重要因素。因此,本研究中进一步分析了ZmNF-YC13基因在高温和渗透胁迫下的表达特点。
结果表明,ZmNF-YC13基因受高温和渗透胁迫快速诱导表达,说明ZmNF-YC13可能是一个响应热胁迫应答的转录调控因子。
然而,提高ZmNF-YC13基因的表达量虽能提高植株对内质网胁迫的耐受能力,但过高的ZmNF-YC13表达量同时也抑制植株叶片的生长。
为了能利用提高ZmNF-YC13基因表达量,来增强植株的抗逆性,同时又尽可能消除过高的ZmNF-YC13表达量带来的不良效应,我们创制了利用拟南芥热胁迫应答基因AtHSP70的启动子,在玉米中驱动ZmNF-YC13基因表达的转基因玉米材料。
高温条件下的表型分析也证实了,通过热诱导表达ZmNF-YC13基因,能够增强玉米植株的耐热能力。
本研究结果不仅表明ZmNF-YC13可通过调控HSP70和HsfA2c等基因的表达来响应热胁迫,还揭示了DnaJ类分子伴侣因子也是ZmNF-YC13蛋白调控的重要下游基因。
因此,本研究不仅拓展了NF-YC转录因子在热胁迫通路中调控的下游基因,同时也证实了植物内质网胁迫应答和植物热胁迫应答在调控通路上存在交叉。
本研究结果表明,玉米核因子编码基因ZmNF-YC13受高温和渗透胁迫诱导表达,在玉米中,热诱导表达ZmNF-YC13可提高玉米苗期植株的耐热能力。
对野生型和转基因材料在高温条件下的表达分析表明,ZmNF-YC13在热胁迫条件下可增强热胁迫应答基因的表达。
荧光素酶报告基因实验和ChIP-qPCR实验表明,ZmNF-YC13还能调控热激转录因子编码基因ZmHsfA2c的表达。
因此,本研究结果表明,ZmNF-YC13是玉米响应热胁迫应答过程中的重要调控因子,热诱导表达ZmNF-YC13可能是通过增强下游热胁迫应答基因的表达,来提高植株的耐热能力。









